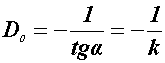 ,
,A biológiai sugáreffektusokat értelmező másik fontos elképzelés a direkt és indirekt sugárhatás elmélete. A direkt hatásnál a primer fizikai és biológiai folyamat azonos pontban, azonos közegben megy végbe (Sommermeyer, 1938). Az indirekt hatásnál az energiaelnyelés a reagáló biológiai rendszer környezetében megy végbe, a hatás kialakulásához közvetítő közegre, energiaátvitelre van szükség. Ezért nevezzük nem közvetlen, indirekt hatásnak. Az élő sejtekben ilyen alkalmas közvetítő fázis a mindig jelen lévő víz. Indirekt effektus esetében szabad gyökök, gyökionok képződnek, amelyek igen rövid élettartalmúak (kb. 10-5 sec) és nagyon affinisek más fontos molekulákhoz. Az un. reakcióidők igen kicsinyek.
A biológiai anyagok kétfélék: szárazak és nedvesek (vizes oldatok, in vivo állapot)
A száraz anyagokban az ionizáló sugárhatás direkt formája lép fel és az inaktivációhoz nagy dózisok szükségesek. A nedves anyagokban mind a direkt, mind az indirekt hatás jelentkezhet, s az inaktivációs dózisok kisebbek. Élő rendszerekben a direkt és indirekt sugárhatást számos tényező befolyásolja (l. később a biofizikai változások és hatások tárgyalásánál!), (Donhoffer, Várterész, Alexander és Bacq, Herrmann).
A direkt és indirekt hatásmechanizmus elmélete az un. találatelméletre épül, amely feltételezi, hogy: a biológiai hatáshoz vezető primer fizikai folyamat, a sugárenergia elnyelődése egy jól definiálható helyen: a sugárérzékeny térfogatban megy végbe, e térfogatban (target, Zielvolumen) tekintélyes mennyiségű energia szabadul fel (hit, Treffer), amelyet találatnak nevezünk (Crowther, 1924, Lea, 1946, Korogodin), az összes sejt sugárrezisztenciája azonos és a sugáreffektus független a hőmérséklettől.
A találat-elmélet (target theory, quantum hit theory, direct effect theory, Trefferprinzip) rendkívül hatékonynak bizonyult a radiobiológiában, jóllehet nem ad mindenre magyarázatot. Jelentős fejlődésen ment át, elegáns matematikai eszköztárat épített fel (Dessauer, 1922, Sommermeyer, 1940, Timoféeff-Ressowsky, 1942, Korogodin, 1966, Dertinger és Jung, Kiefer és Koelzer). Mint azt korábban már említettem, a kérdés nagy ívű összefoglalását Lea adta meg idézett könyvében, s a magyar nyelvű források is jelentősek (Kovács P., Köteles, Rontó, Sztanyik, Tarján).
Mivel a találat-elmélet számos hiányosságára fény derült, a kutatók különböző módosításokkal próbálkoztak (pl. Jordan-féle felerősítési, un. amplifier, vagy Verstärker elmélet, 1937, 1938, a kémiai mérgezés, vagy indirekt hatás elmélete, az angol nyelvű forrásokban “poison theory"-nak nevezett modell, a Minder-féle fiziko-kémiai elmélet 1946-ból, a biológiai sugárhatás matematikai elmélete, amelyet Blau és Altenburger irt le 1922-ben, a histamin-elmélet Ellinger 1951-ben közölt dolgozatában, stb.).
A találat-elmélet fogyatékosságai ellenére alkalmas arra, hogy a biológiai sugáreffektus értelmezésében a direkt sugárhatást magyarázni tudjuk vele. Segítségünkre van ebben a jelenségek grafikus, szemi-logaritmikus ábrázolási módja, az un. dózishatásgörbék alkalmazása (4. sz. ábra).
A direkt hatás okozta sugárbiológiai változásokat két görbetípus írja le: az exponenciális és a nem exponenciális dózishatásgörbe (dose effect curve).
Az exponenciális görbéknél a találatelmélet leegyszerűsített formájában feltételezi, hogy a sugárérzékeny objektumok túlélő hányada a dózissal exponenciálisan csökken. Más szavakkal: az inaktivációhoz csak egy találatra van szükség (single hit). Az egy találat fogalom bevezetése a radiobiológiába Henry nevéhez fűződik (1905). Az ilyen görbéket nevezzük egytalálatos görbéknek (single hit curves). A találatelmélet révén néhány nem exponenciális természetű dózishatásgörbe szintén értelmezhetővé vált.
A nem exponenciális görbéknek további két típusa van: a több céltárgyas modell (multi target curves), ahol a biológiai hatás eléréséhez a biológiai objektum "n" számú sugárérzékeny térfogatának (target, Zielvolumen) mindegyikét inaktiválni kell, a másik az un. többtalálatos görbetípus (multihit curves) - ebben az esetben a biológiai céltárgyak minden esetben egy "target"-nek felelnek meg, de ezek inaktivációjához több találatra (hit, Treffer) van szükség.
Általános gyakorlat, hogy a mérhető sugáreredetű eltérés (pl. túlélés) valószínűségének logaritmusát a dózis függvényében rendszerint féllogaritmusos koordinátarendszerben ábrázolják. A kapott dózishatásgörbéket konvencionális paraméterekkel jellemezhetjük, akár normális, akár neoplasticus sejtekről legyen szó. A 4. sz. ábrán egy nem exponenciális, un. vállas-görbe (shoulder curve) látható. A D1-érték a görbe kezdeti lejtésének (iránytangensének) inverze, a D37-érték pedig a 37%-os dózistúlélést jelzi. Ez utóbbi adat - nemzetközi megállapodás alapján - az e-1 = 0.37 értékből adódik, ahol az e-paraméter a természetes logaritmus alapszáma (Cavete! D37 = LD63!). A D0-paraméter a dózishatásgörbe magasabb dózistartományban exponenciálissá (a semilogaritmusos koordinátarendszerben egyenessé) váló görbe iránytangensének negatív reciproka. Azaz:
,ahol a k-érték a lejtésszög tangensének szimbóluma (Kovács P., 1971). Feltételezik, hogy az exponenciális szakasz meghosszabbítása a log-skálán egy olyan értéket metsz ki (n-szám, vagy extrapolációs szám), amely a szükséges sugárérzékeny térfogatszámmal, vagy találat-számmal lesz azonos. Az n-szám értéke normális emlős sejteknél 2-10 közé esik, daganatos sejteknél ez több. Mivel időközben kiderült, hogy az n-szám értéke sejttípus és sejtciklus-fázis függő, ma úgy látják, hogy helyesebb ezt az értéket extrapolációs számnak nevezni, semmint sugárérzékeny céltérfogat-, illetve találat-számnak (Fowler). A három D-paraméter csak minősített esetben lesz (lehet) azonos értékű egymással, ha teljesül az n=1 feltétel, ha a görbe exponenciális (egytalálatos, egyenes lefutású). Amennyiben nem ez a helyzet (ną 1!), úgy az alábbi egyenlőtlenség az igaz:
Az elmélet szerint a nem exponenciális görbék vállszakasza a biológiai objektumok sugárhatás utáni rezisztenciáját, illetve regeneratív kapacitását jellemzi. E váll minél nagyobb, annál nagyobb a rezisztencia, annál sikeresebb a sugársérüléseket kijavító mechanizmus. A vállhossz mérőszáma a Dq-érték, amelynek külön neve is van: “quasi treshold dose" (Alper et al., 1962). Az emlős sejtek Dq-paramétere in vitro ~2-3 Gy között mozog, in vivo elérheti az 5Gy-t (Fowler, i.m., 1962). A dózishatásgörbén grafikusan úgy adhatjuk meg a Dq-értéket, hogy az exponenciális szakasz meghosszabbítását a 100%-os túlélés magasságában, az abszcisszával paralell, metsszük. A metszéspontot a dózistengelyre levetítve leolvashatjuk a megfelelő értéket. Az ismertetett görbeparaméterekről tudni kell, hogy a dózishatásnak nem abszolút értékei, másfelől nemcsak a sugárzásokra, hanem a cytostaticus hatásra is alkalmazhatók.
A klasszikus találatelmélet szerint a D0-érték azt a röntgensugárdózist jelenti, amely szükséges ahhoz, hogy valamennyi sugárérzékeny térfogatban egy letális találat jöjjön létre. A D0-érték emlős haemopoeticus őssejteknél (stem cell) 0.83-1.15 Gy között változik, élettani körülmények között, egyéb sejtfajtáknál és in vitro kb. 51-480 rad (0.51-4.80 Gy) közé esik (Willmer). Cytostaticumok hatásánál a D0-paraméter 0.02-200.0 µg/ml, az n-szám: 1.2-8.0 (Mauro és Madoc-Jones, 1970). A két érték szőrtüszőknél 1.8-2.2 Gy, ill. (az n-szám) kb. 6 (Potten, 1968), ugyanez a bélhámsejteknél: D0~0.97 Gy, n~35-160 (Withers és Elkind, 1968). Ez utóbbiaknál, a tíznél nagyobb extrapolációs szám nem meglepő, hiszen a bélhámsejtek proliferációs sebessége nagy, a ciklusidő kicsi, a populációk ciklusbeli eloszlása heterogén (aszinkron). Porcsejteknél a D0~1.9 Gy, az n-szám ~ 6 (Kember, 1965).
Malignus szövetekben in vitro a D0~1.09-1.62 Gy, az extrapolációs érték ~ 1.6-10.0 közé esik.
Általában igaz, hogy a többtalálatos görbéknél a D0-paraméter értéke a sugárfajta (RBW, LET!), a dózisteljesítmény, a frakcionálás, az oxigén koncentráció függvényében változik. Az egytalálatos görbéknél mindezektől független. Magas ionizációs sűrűség esetében a vállszakasz eltűnik, Dq-érték nem képződik, n=1, a görbe egyenes lefutásúvá válik.
A dózishatásgörbék elméletét és e görbék konvencionális paramétereit már ma is mindannyiunknak ismernie kell, mivel alkalmazásuk a klinikai gyakorlatban és a sugárvédelemben egyre terjed (Fliedner, 1995).
A vállas túlélési görbék a találatelmélet alapján a következő egyszerű függvénnyel, exponenciális egyenlettel írhatók le:
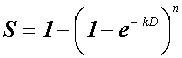 ,
,ahol: S (survival) = a D dózist túlélő frakcióval, az e a természetes logaritmus alapszáma, a k az exponenciális görbeszakasz meredekségét jellemző szám, az n az extrapolációs érték (Lea, 1946).
Az indirekt hatásnál a biológiai következményekért a szabad gyökök, a gyökionok, a gyökkomplexek felelősek. Korábbi véleményekkel szemben ma egyre többen elfogadják, hogy az indirekt hatás nemcsak vizes rendszerekben, hanem száraz anyagokban is létezik. A jelenség legjobban ismert modellje a víz radiolysise, ahol igen diffusibilis és reaktív OH-, H-, eaqu és H2O2-gyökök képződnek. Radiobiológiai jelentőségük abban van, hogy mintegy “felerősítik" a primer energiaelnyelődés elemi sugárhatását. Az említett szabad gyököknél kiderült az a lehetőség is, hogy kedvező feltételek esetén ismét vízmolekulává rekombinálódhatnak. Mai eszközeinkkel a direkt és indirekt hatást egymástól még nem tudjuk elkülöníteni. A Dale-féle hígítási effektus azt igazolja, hogy a direkt hatás esetén a növekvő hígítás csökkenti a sugárhatás mértékét. A hőmérséklet hatásával szemben a direkt sugárhatás kevésbé érzékeny, mint az indirekt (Herrmann, 1990).