 |
2-1. Az “elemi sugárhatás"
A korábbi előadásokon már elhangzott, hogy fizikai szempontból a hullámtermészetű és korpuszkuláris ionizáló sugárzásokat különböző energia formáknak kell tekintenünk. Másfelől: a sugárbiológiai hatás kialakulásában a primer effektus a sugárenergia elnyelődése az élő anyagban.
Az elemi sugárhatás összefoglaló neve mindazon történéseknek, amelyek a primer fizikai folyamat, a sugárenergia elnyelődése és a biológiai válasz közti időszakban lépnek fel. A radiobiológia klasszikus tanítása szerint az elemi sugárhatás több lépcsőben megy végbe. Ennek szakaszai a következők: fizikai, fiziko-kémiai, kémiai - biokémiai és biológiai szakasz.
Míg a fizikai fázisban nano- és milliszekundum nagyságú időtartamokkal számolnak, a következő lépéseknél az idő növekszik. A biológiai szakaszban, pl. perc, óra, nap, hetek, évek lesz (lehet) a lépték. A primer folyamat, az energiaelnyelődésének értelmezése két pilléren nyugszik: a kvantum- és az atom-elméleten.
Az abszorpció fizikai formái a következők: excitatio, fotoelektromos abszorpció, Compton-effektus, párképződés, nukleáris fotóeffektus és elasztikus ütközés (recoil-proton kiválás).
Az említett kölcsönhatástípusok meghatározott sugárenergiákhoz kötöttek. Ahogy nő az elnyelt sugárzás energiája, úgy haladunk az excitatio utáni jelenségek irányába. A klinikai sugárbiológiában elsősorban olyan energiatartományokról van szó, amelyeknél a fotoelektromos abszorpció, a Compton-effektus és ritkábban a párképzés a domináns elnyelődési forma. Az elnyelt sugárenergia esszenciálisan két domináns fizikai jelenséget idéz elő: az atomok és molekulák ionizációját, illetve azok gerjesztését (excitatio). Az ionizáció során az ionpárok képződése különböző módon történik (fotoelektromos abszorpció, Compton-effektus, párképzés) és a biológiai hatás a képződött másodlagos elektronokra vezethető vissza. Megjegyezzük továbbá, hogy csak az elnyelt sugárenergia fejt ki radiobiológiai hatást (l. később a Grotthus-Draper törvénynél!).
A fizikai hatás az energiát elnyelő anyagban (közegben) olyan közös egységet hoz létre, amelynek összetevői az: exitált elektronok (plasmonok), az ionok és a másodlagos elektronok (delta-sugárzás).
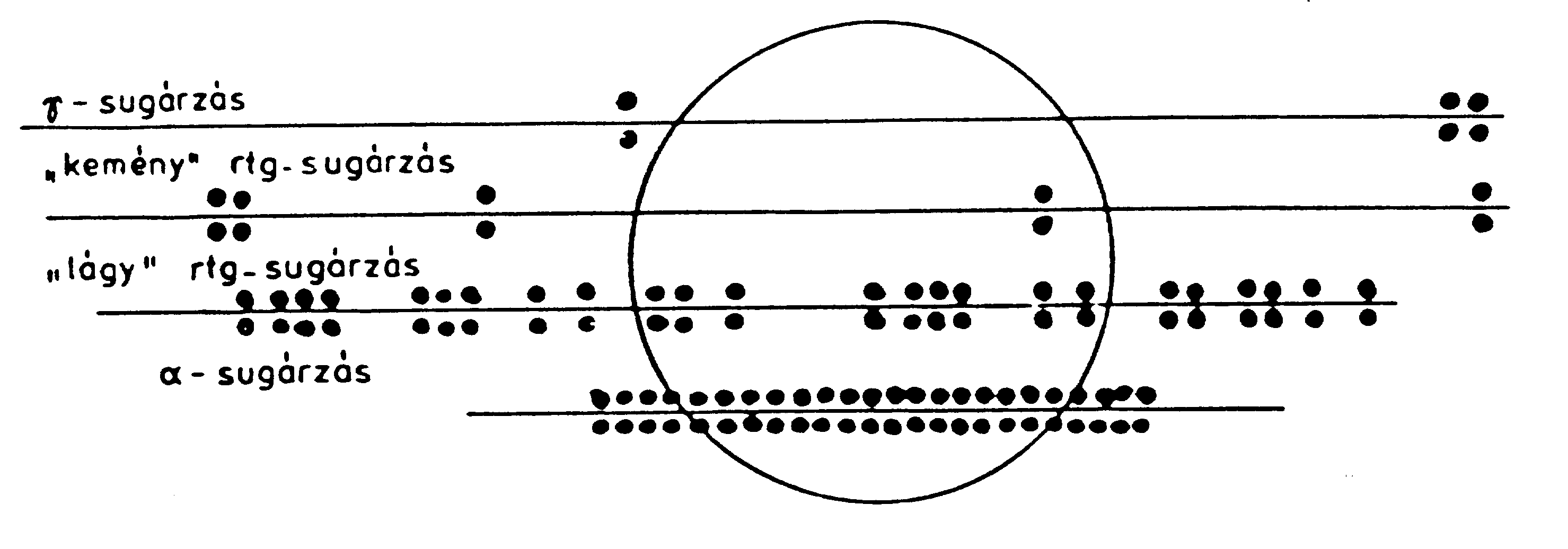
Ezt a közös fizikai egységet ion-, vagy elektron-felhőnek (cluster) is nevezik. A primer töltött részecskék és másodlagos elektronok, mozgásuk során és útvonalaik mentén, gerjesztett és ionizált molekulákból álló, bonyolult szerkezetű ion- és elektronsűrűségeket képeznek, amelyek térbeli eloszlása és társulásai igen összetettek. Az egyes részecskék pályájukon egymás utáni, sorozatos ütközéseket (collisions) szenvednek el, miközben minden egyes ütközésnél az abból eredő energiavesztés véletlenszerű lesz. Korábban már láttuk, hogy az elnyelt sugárenergia és a másodlagos termékek térbelisége alapvetően fontos a biológiai folyamatok kialakulásában. Az ionsűrűségek pályamenti eloszlása (tracks), a sűrűségszerkezet és a sugárminőség (LET!) az akut és kései sugárhatásokban döntő tényező. A közvetlenül és közvetve ionizáló sugárzások lineáris ionsűrűségeit, az energia (penetráció) függvényében a 3. sz. ábra (Fritz-Niggli, 1970.) modellezi.
|
A kérdés első, máig nélkülözhetetlen összefoglalását Lea (1946.) írta meg. Nézete szerint az elemi sugárhatásban létrejött ionsűrűség-eloszlások (makrorendszer) a pályamentén véletlenszerűen eloszló közös fizikai egységek, az ion- és elektronfelhők alrendszereiből épülnek fel (mikrorendszer). Azt, hogy az ionsűrűségek térbeli eloszlása és ezek egymáshoz való kölcsönhatása a biológiai hatásban nagyon fontos, 1930-tól ismerjük. További fejlődést eredményezett az elemi sugárhatás vizsgálatában, az 1970-ben bevezetett Monte-Carlo - módszer (Goodhead, 1995). A szakirodalom bőséges volta azonban jelzi, hogy még nagyon sok a megoldatlan feladat! Számos kiválóság és nagy egyéniség ért el e területen maradandó eredményeket (Dessauer, 1923., Holthusen, 1928., Timoféeff-Ressowsky, 1934., Failla, 1936., Lea, 1946., Rajewsky, 1956., Bacq és Alexander, 1961., Várterész, 1963., Hug és Kellerer, 1972., Fritz-Niggli, 1973., Korogodin, 1975., Herrmann, 1990.). E témacsoport kiemelt helyet kapott a Röntgencentenárium alkalmából rendezett világkongresszuson is (Radiation Research 1895-1995 - 10. ICRR '95 - 100 years X-rays - Tenth International Congress of Radiation Research, Würzburg, Germany, August 27 - September 1., 1995.).
Az elemi sugárhatás folyamatában a fizikai és a biológiai szakasz között helyezkedik el a közbülső, a fiziko-kémiai és kémiai-biokémiai lépcső. Az elnyelt sugárenergia kiváltotta kémiai reakciók fázisa ez, sok forrás a fiziko-kémiai és kémiai-biokémiai szakaszt csupán kémiainak nevezi. A kémiai sugárhatást több elmélet próbálta megmagyarázni. Néhány ezek közül: termokémiai, vagy ponthő-elmélet (point heat theory, Punktwärmetheorie, Dessauer, 1923.) - ez feltételezi, hogy az energia-elnyelődését követő excitatios és ionizációs energia kinetikus impulzussá alakulván a molekulák diszkrét pontjaiban pontszerű, elemi fizikai hőhatást fejt ki, ez vezet a kémiai effektushoz; sugárkémiai elmélet /Holthusen, 1928./ - az elnyelt sugárenergia, az anyag excitatioja és ionizációja magasabb szintű energiaállapothoz vezet, amely normális állapotú molekulákban nem észlelhető kémiai reakciókat tesz lehetővé (radiochemical theory); fiziko-kémiai, vagy folyadékáramlási elmélet (fluid flow theory, Failla, 1936.) - az energia-abszorpciót követő gerjesztés a kiindulási molekulánál (anyagnál) alacsonyabb molekulasúlyú substantia képződéshez vezet, amelynek következménye az intracellularis ozmotikus nyomás változása lesz; kolloidkémiai elmélet (colloidal-chemical theory, Hardy, 1903.) - az élő anyagban kolloid-oldat rendszerek vannak jelen, amelyben pozitív vagy negatív töltések szuszpenziója is kimutatható. Az elmélet feltételezi, hogy a sugárhatás e rendszerek töltését változtatja meg, aminek a következménye a kolloid-stabilitás változása.
A kémiai sugárhatás vizsgálata során kiderült, hogy az effektus kialakulásában két folyamat, a radiokémiai és fotokémiai reakció a vezető történés (ionizáló és nem ionizáló sugárzások hatásakor). Ellinger (1957) véleménye szerint a sugárhatás rádio- és fotokémiai elmélete a jelenségek többségét jól magyarázza. Mivel az élő anyag és a vizes oldaltok kapcsolata nyilvánvaló, vizsgálni kell az oldott anyagok, az oldószer, a koncentráció, a hőmérséklet és a kísérő anyagok szerepét a sugárkémiai folyamatokban, a változások hatásfokát. Jobban meg kell ismernünk a vízre (szabad gyökök), a proteinekre (denaturatio, kémiai reakció, flocculatio), az aminosavakra, az - SH-vegyületekre, a nukleinsavakra, a zsírokra, a szénhidrátokra, a hormonokra, a vitaminokra, az enzimekre és mindezek elegyére (kevert hatás, dilutios effektus) gyakorlott sugárhatást. A kémiai-biokémiai reakciókat az egyes szakkönyvek korábban csak fejezetnyi terjedelemben tárgyalták. Az első modern radio-biokémiai könyv 1969-ben jelent meg (Streffer, 1969).
Az elemi sugárhatás utolsó fázisa a biológiai reakciók jelentkezése. Az élő anyagra kifejtett általános sugárhatás térbeli eloszlása alapján lokális és általános, hatásmechanizmusa szerint direkt és indirekt, az időfüggését tekintve akut és kései, minőségét illetően morfológiai és funkcionális, illetve mennyiségileg a reparatiot tekintve reversibilis, feltételesen reversibilis és irreversibilis, a lefolyását tekintve pedig lépcsőzetes és minden, vagy semmi típusú lehet (graded action type, Clark, 1937., all or nothing type).
A biológiai szakaszban jelentkező alapelváltozások három csoportra oszthatók: a morfológiai, a funkcionális és az általános sejtélettani következmények csoportjára.
A post irradiationem (p.i.) fellépő sejtalaktani reakciók mind a sejtmag, mind a cytoplasma részéről általánosan ismertek. A mag sugársérülésekor a chromatin állomány kisebb fokú károsodása mellett (vacuolisatio, pyknosis, kromoszóma-szerkezet sérülése) a durvább ártalom is kialakulhat (karyorrhexis). A plazma károsodásának leggyakoribb jele a vacuolisatio, de ugyanígy sérülhetnek a sejtorganellumok (pl. a mitochondriumok, Golgi-apparatus, stb.) is. Nem tisztázott máig sem, hogy a plasmogének, a nukleinsav-tartalmú képletek sugárártalma mennyiben befolyásolhatja a radiobiológiai effektust. A sejtplazma korábban nagyobbnak vélt sugárrezisztenciája csak látszólagos!
A funkcionális változások közül a sejtmobilitásra, a növekedésre, az anyagcserére, a sejtpermeabilitásra, a reprodukcióra kifejtett sugárhatás a fontosabb.
Az általános sejtbiológiai effektusok közül is kettőt említünk meg: a sugárzás okozta sejthalált és a stimulációt.
A sugárenergia elnyelődése a sejt elpusztulását okozhatja közvetlen destrukcióval (koagulációs nekrózis, cytolysis), letális mutáció és korai elöregedés révén. Újabban ide soroljuk még a programozott sejthalál, az apoptosis lehetőségét is. Heves vitát váltott ki a radiogen stimuláció feltételezése. Az ötlet nem új (Maldiney és Thonwenin 1896-ban, növényekben sugárhatás után gyorsult csirázást észlelt), és jelenleg a Lucky-féle hormesis-elméletben él tovább. Emlős és humán sejtekben nincs azonban ez utóbbira meggyőző bizonyíték. Az ionizáló sugárzások hatását bionegativnak tekintjük továbbra is. A radiológiatörténet későbbi szakaszában keletkezett, sugár-hatást magyarázó elméletek alapját a bionegativitás ténye képezte és képezi máig. Ezek az elméletek tehát feltételezik és elfogadják kiindulási alapként, hogy: az ionizáló sugárzás bármilyen mennyisége bionegativ hatású, az enigma, a stigma megmarad, a sugárhatást az élő rendszerek nem “felejtik el" (Kelen, 1928., Ratkóczy, 1959., Zsebők, 1979.) és a sugárbiológiában nem érvényes a gyógyszertanból és toxikológiából ismert Arndt-Schulz törvény (ez utóbbi szerint: kis dózis stimulációt, közepes depressziót, a nagy pedig destrukciót okoz), nem általános érvényű Paracelsus megállapítása: "... alleine die Dosis macht, wenn ein Ding Gift ist, oder nicht".
A hormesist növény-modellben írták le, amely egészen más sugárbiológiai rendszer, mint az emlős és emberi sejtek és szövetek rendszere. Az állati és humán sejtekben és szövetekben leirt látszólagos “serkentő hatás" nem tekinthető biopozitívnak, tehát hormesisnek. Elsősorban azért, mert nagy valószínűséggel egy adott morfológiai (anatómiai) struktúrához kötött élettani-biokémiai folyamatpárok egyikének depressziója a jelenség háttere, a másik antagonista viszonylagos túlsúlyával. (Bond, 1995, Fliedner, 1995, Bebesko, 1995, Trott, 1997). A kompetitív antagonizmus egyensúlyának felborulásáról van szó.